A vadalma (Malus sylvestris L.) génmegőrzése
Nyári László
NyME Környezettudományi
Intézet
A vadalma (Malus sylvestris
(L.) Mill.) genetikai identitását veszélyeztető okok
E ritka erdei gyümölcsfajunk
szálanként fordul elő, példányai egymástól izolálódtak. A megritkulás okát elsősorban
előhelyének beszűkülésében kereshetjük. A vadalma, de a történelmi, hazai alma
gyümölcsfajták termőhelye az időszakos elárasztást kapó – időszakos vízhatású
ártéri termőhelyek – az ármentesítés következtében mind csökkenő mértékben irhatóak
le ennek megfelelő termőhelytípus-változatként, ill. e tarsulásokat sok helyütt
más kultúrtársulásokkal váltották fel.
Kleinschmit,
J, Wagner, I. (1995) három fő szempontot emelnek ki a megritkulás okaiként:
1.) A gazdasági érdekeltség
hiánya. A természeti viszonyokhoz igazodó gazdálkodás idején e fajok – akárcsak
napjaink hazai erdőgazdálkodásában – gyümölcsükért voltak leginkább ápolva.
A vadalmát mint marhaeleséget is számon tartották, mindamellett, hogy vadtakarmányként
is számoltak vele, de az emberi táplálkozásban is szerepet kapott. Andrásfalvy
B. (1976) részletesen közli a Duna Tolna megyei szakaszán megfigyelt
tartósítási – aszalási, ill. a piacra szálllítás szokásait, mely igazolja –
kb. a második világháború végéig tartó – un. ártéri gyümölcsgazdálkodás meglétét
még a század első felében is. Természetesen e gazdálkodási mód nem a vadalmát
részesítette kifejezett előnyben.
2.) Megváltozó erdészeti
célkitűzések, melyek a sarj- és középerdőket szálerdővé alakítva egészen más
konkurenciaviszonyokat eredményeztek az erdei vadgyümölcsök számára; mindamellett,
hogy egyes szakirodalmi források "erdészeti jelentőség nélkülieknek" minősítve
őket, nem járultak hozzá további megbecsülésükhöz.
3.) A mezőgazdálkodás
fokozódó belterjessé válása, monokultúrák kialakulása, melynek révén a patakok
mentének erdőtársulásait megritkították, a kisebb vadbúvó csendereseket, facsoportokat
kiirtották, ami a területek faji diverzitásának degradalódásához, a terület
faji lefedettségének csökkenéséhez vezetett. Pl.: Buttenschon, R. M, Buttenschon,
J. (1998) Dánia egy mozaikszerű erdőfoltokkal tarkított legelőterületén
vizsgálta egy 1800 egyedet számláló magoncpopuláció sorsát. Az egyedek 98 %-ánál
elsősorban lovak kisebb mértékben marhák általi rágáskár volt megfigyelhető.
Az "újulat" megmaradása főként a legeltetés intenzitásától függ. Várhatóan 20
%-uk maradna életben, ha a csírázás utáni első esztendőkben szüneteltetnék vagy
mérsékelnék a legeltetést, ellenkező esetben a túlélők száma minimális.
Az erdei vadgyümölcsök,
tágabb értelemben a nemeslomb elegyfafajok iránti fokozottabb érdeklődést e
fafajok faanyaga iránt mutatkozó kereslet, és az így elérhető magasabb árbevétel
is nagyban indukálja. Az erdőgazdák figyelme tehát a nemeslomb elegy (noble
hardwoods) iránt nem teljesen önzetlen, és nem csupán szakmai elkötelezettségből
táplálkozik. Ez a folyamat azonban esélyt ad arra, hogy e megritkult, genetikai
állományában a leromlás fenyegetettségének, ill. a genetikai identitásában veszélynek
kitett fajok helyzetét egy ökológiai szemléletű erdőgazdálkodásban biztosíthassuk
(Schuman, K, 1989; Kleinschmit, J, 1998).
A faj egyedszámának
megritkulása mellett és azzal egyetemben jelentkezik fokozódó veszélyként a
genetikai szennyeződés fogalma. Mivel a vadkörte és vadalma meghatarozó mertékben
nem önbeporzó fafajok (Janb en, A; Walter, P, 1997), ezért
az egymástól elszigetelt, elöregedő szálankénti előfordulások gyakorlatilag
a rendelkezésre álló felkultúr ill. kultúrfajtákkal basztardizálódnak. Az így,
ivaros szaporodás révén keletkező utódnemzedék nem az eredeti fajra jellemző
genetikai információt viszi tovább.

4. sz. kép: Az R 60-as törzsfa Escherode-ban nevelt utódnemzedékének
termései
(Kleinschmit, J., Wagner I., 1995) |
Német kutatók közlése szerint (Kleinschmit,
J., Wagner I., 1995) egy szálankénti előfordulásban található
egyed generatív utódainak vizsgálata során a termésalak, de különösen
a méret és a szín jelentős különbözőségeket mutatnak, melyek meghaladják
a faji azonosság morfológiai tűréshatárait (4. sz. kép). Itt a legnagyobb
alma egy erős fokú, kultúrfajtákkal történt hibridizációra utal, míg egy
másik utód cseresznye nagyságú termésében egy kis termésű díszalma genetikai
befolyását figyelhetjük meg.
|
A hibrid eredetű
Malus domestica általában ritkábban kereszteződik, nem vesz részt a nagyszámú
hibrid eredetû díszalma léterhozásában. A Malus domestica nem ellenálló,
a faj kivadult egyedei általában nem jutnak el a virágzásig. A Malus pumilla
ill. ennek szibériai alfaja a Malus pumilla 'Niedzwetzkyana' vesz részt
a díszalmahibridek képzésében, amely hibridek intenzívebben kereszteződnek
(Fr. Fiala, J. L, 1994).
Felmerül természetesen
a kérdés, hogy ez a fajta genetikai szennyeződés mikor jelentkezhetett elsőként,
mekkora volt a kereszteződéseknek, ill. az arra való hajlandóságnak a "genetikai
nyomása". Egy sűrűbb, kiterjedtebb lefedettség – mely a saját pollen genetikai
áramlását is biztosította – mennyire ellensúlyozta ezt a folyamatot. A középkori
Magyarország területén is fejlett gyümölcskultúra virágzott, melynek hatását
figyelembe kell vennünk. Vancsura R. (1992, ex verbis 1998) közlése
szerint a nemes alma fajok a vadalmával rendszerint nem kereszteződnek. Az FM
NGT Erdészeti Munkabizottságának jelentése alapján nem bizonyított, hogy a vadalma
a nemes alma fajtáival kereszteződik (Bartha D, Bordács S, 1998).
A vadalma társulásai
Európai (mediterrán)
flóraelem, planicien-kollin-montán vertikális, ill. Közép-, Nyugat- és Dél-Európára
vonatkozó horizontális elterjedésû fafaj. A Tiszántúl kivételével hazánkban
közönséges. Ökológiai igényeit illetően Ellenberg a fényigényességet említi
(in.: Wagner, I, 1995); Vancsura R. (1992) félárnyékűrésről
ír. Fagytűrő képességét illetően megoszlanak a vélemények. Társulási optimumát
az erdő nedvességhatáránál találjuk, bár dombvidékeinken és középhegységeink
melegebb oldalain is előfordul. Az üde, tápanyagban gazdag termőhelyeken gyakori.
A meredek fekvésű, főként déli és nyugati kitettségű szárazabb erdőkben társulási
viszonyai nagyrészt a vadkörtével megegyeznek (Hoffmann, H, 1993).
de ezen növényszövetkezetekben jelentéktelenebb szerepet játsszik, jóllehet
a laza záródású koronaszint kedvezőbb fényellátottsági viszonyokat jelent a
vadalma egyedek számára is.
Az alábbiakban közlöm
azokat a vadalmával többé-kevésbé jellemezhető erdőtársulások listáját, melyet
Bartha D. előzéke, valamint Csapody I., Csapody V., Rott
F. (1966) és Bartha D., Kevey B., Morschauser T., Pócs T. (1995)
alapján, ill. saját megfigyelésekre építve állítottam össze:
3. sz. táblázat.
A vadalmával jellemezhető erdőtársulások listája.
|
Keményfás ligeterdők (1)
|
|
Tölgy - kőris - szil liget
|
Fraxino pannonicae - Ulmetum
|
|
Égerliget
|
Paridi quadrifoliae - Alnetum
|
|
Patakmenti ligeterdők - Domb- és hegyvidéki ligeterdők
(1)
|
|
Hegyvidéki égerliget
|
Carici brizoidis - Alnetum
|
|
Gyertyános égerliget
|
Aegopodio - Alnetum
|
|
Szubmontán sásos égerliget
|
Carici acutiformis - Alnetum
|
|
Hegyvidéki kőrisliget
|
Carici remotae - Fraxinetum
|
|
Gyertyános - tölgyesek (2)
|
|
Dél-dunántúli gyertyános - tölgyes
|
Helleboro dumetorum - Carpinetum
|
|
Gyertyános - kocsányos tölgyes
|
Querco robori - Carpinetum
|
|
Dél-dunántúli gyertyános - kocsányos tölgyes
|
Fraxino pannonicae - Carpinetum
|
|
Homoki tölgyesek (2)
|
|
Nyílt homoki tölgyes
|
Festuco rupicolae - Quercetum roboris
|
|
Zárt homoki tölgyes
|
Convallario - Quercetum roboris
|
|
Lösztölgyesek (2)
|
|
Lösztölgyes
|
Aceri tatarico - Quercetum
|
|
Meleg- és mészkedvelő tölgyesek (2)
|
|
Melegkedvelő tölgyes (Molyhos - kocsánytalan tölgyes)
|
Corno - Quercetum (pubescenti - petraeae)
|
|
Dolomittölgyes
|
Cirsio pannonici - Quercetum pubescenti - petraeae
|
|
Nyúlfarkfűves tölgyes
|
Seslerio - Quercetum virgilianae
|
|
Magyarperjés andezittölgyes
|
Poa pannonicae - Quercetum petraeae
|
|
Mészkedvelő tölgyes (Molyhos - cseres tölgyes)
|
Orno - Quercetum (pubescenti - cerris)
|
|
Mecseki mészkedvelő tölgyes
|
Rusco - Orno - Quercetum
|
|
Sziki erdők (2)
|
|
Sziki tölgyesek
|
Festuco pseudovinae - Quercetum roboris
|
|
Sziklaerdők (3)
|
|
|
Hárs - kőris sziklaerdő
|
Tilio - Fraxinetum
|
|
Cseres - tölgyesek (3)
|
|
Cseres - kocsánytalan tölgyes
|
Quercetum petraeae - cerris
|
|
Mezei juharos - tölgyes
|
Aceri campestri - Quercetum petraeae - roboris
|
|
Hársas - tölgyes
|
Dictamno - Tilietum cordatae
|
|
Ezüsthársas - cseres - kocsánytalan tölgyes
|
Tilio tomentosae - Quercetum petraeae - cerris
|
|
Cseres - kocsányos tölgyes
|
Quercetum robori - cerris
|
|
Bokorerdők (3)
|
|
Sajmeggy - molyhos tölgy bokorerdő bazalton és mészkövön
|
Ceraso (mahaleb) - Quercetum pubescentis
|
|
Sajmeggy - molyhos tölgy bokorerdő andeziten
|
Festuco pseudodalmaticae - Ceraso - Quercetum
|
|
Cserszömörce - molyhos tölgy bokorerdő
|
Cotino - Quercetum pubescentis
|
|
Baranyai peremizses karsztbokorerdő
|
Inulo spiraeifolio - Quercetum pubescentis
|
Jelmagyarázat:
(1): gyakori; (2): közepes; (3):
ritka előfordulás.
A vadalama erdőnevelése
Az erdőnevelés folyamán
elsősorban a magonc eredetű újulatra számíthatunk, mivel a vadalma rosszul sarjadzik,
és a szórványosan megjelenő tősarjak későbbi fejlődése gyenge (Vancsura
R, 1992).
A vadalma a vékonyrudas
kor elején még jól bírja a magassági növekedési versenyt, majd később alászorul;
ekkor kap jelentőséget a fényigény kielégítése. Egy a génmegőrzést is szolgáló
erdőgazdálkodásban a befejező tisztítás ill. a törzskiválasztó gyérítés a vadalmát
segítő, kímélő elvégzése nyújthat lehetőséget a fafaj további, állományban történő
megtartására.
A nedves, időszakos
vízhatasú jó tápanyagellátottságú termőhelyekre vonatkozóan további megfigyelések
szükségesek, hogy ezen viszonyok között a vadalma milyen mértékben tudja viselni
a tartós árnyalást – valószínűsíthetően jobban.
A vadalama génmegőrzése
A feladat megfogalmazásakor
legfontosabb kérdés marad, hogy a genetikai állományában fajazonos; nem hibridizálódott,
illetőleg ne a kivadult kultúregyedek genetikai információját tartsuk fenn egyfajta
mesterséges referenciapopulációk, azaz klóngyűjtemények, géngyűjtemények stb.
létrehozásával. Továbbra is vita tárgyát képezheti, hogy a kérdéses esetekben
kivadulásról, avagy kultúrfajtákkal történt hibridizációról van-e szó? E kérdésre
egzakt válasz genetikai vizsgálatokkal (izoenzim-analízis, RAPD, PCR-RFLP) kereshető.
A morfológiai ismérvek
alapján vadalmaként meghatározott egyedeket vegetatív úton továbbszaporítjuk.
Autovegetatív szaporítás
esetében a dugványozas nem kecsegtet kielegítő eredménnyel, kipróbálandó lehetőségként
jelentkezhet az in vitro (mikro)szaporítás alkalmazása. Ez utóbbira vonatkozólag
már vannak tapasztalatok (Janb en, A; Walter, P, 1997).
Heterovegetatív
szaporítás esetében legjobb eredménnyel az un. chip-szemzés alkalmazása mutatkozik
(Kohnert, H, 1991). E módszer lényege azon problémafelvetés megoldásából
táplálkozik, melyszerint az alany és oltógally átmérője jelentősen különbözhet.
A fiziológiailag megújított anyatelepekről persze könnyen nyerhető megfelelő
minőségû oltógally, de az idős példányok, ill. a károsodott reliktum-előfordulások
esetében ez már korántsem ilyen egyszerű szaporítási problémaként jelentkezik.
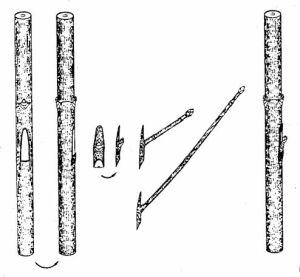
5. sz. kép: A chip-szemzés alkalmazása hegyi juhar heterovegetatív szaporításánál
|
Ezen
szemzési mód végrehajtásakor az oltógallyból egy rügyet (vagy akár vékony
oldalhajtást) egy kis hajtáshéj-pajzsocskával kimetszik, és ezt az alany
megfelelően eltávolított pajzsocskájának helyére illesztik (5. kép). Előnye,
hogy a chip és az alany teljesen pontos illeszkedése nem elengedhetetlen.
Nem szükséges előfeltétel továbbá az alany nedvkeringése; e szemzés nyugalmi
állapotban is elvégezhető. Nagyon jól összenőnek a nagy méretű fás "chipek".
A bél nélküli chipek nem nőnek össze, ilyenkor a fakadás elhúzódik, a növekedés
csekély. Bár e módszert főként hegyi juhar heterovegetatív szaporításánál
használták, vadalmával kapcsolatban is sikeresek a tapasztalatok a Hessen
állambeli Hann. Münden erdészeti kutatóintézetében. |
Az ezen géngyűjtemények
életre hívása során nyert szaporítóanyag szolgáltatja a generatív származású
növényanyagot a repatriálás megvalósításához. Schmidt, J. (1989)
szerint közölt csemetetermesztési technológia az érés és gyû jtés után
megtisztított, magházától megszabadított maganyag késő őszi vetését előnyösebbnek
tartja. Barázdahúzó henger után 1-2 cm mély sorokban fm-ként 1 g magot vetve,
majd azt még vékonyan fűrészporral is takarva, ezáltal könnyítve meg a csíra
kibúvását a tavaszi talajcserepesedés mérséklésével.
Tavaszi vetés esetén
a magvakat előbb szellős helyen óvatosan szárítják, majd decemberben homokba
rétegelik és 2-4 Co-on a vetésig tárolják.
11 % alatti víztartalom,
és 2-10 Co között lezárt konténerekben tárolva a vadalma mag 2 évig
eltartható a csírázóképesség csökkenése nélkül.
Óvatos gyökérmetszés
után szükséges még a magonccsemeték fakadás előtti eliskolázása az életképesebb,több
éves, nagyobb egyedek nevelése érdekében.
Repatriálás
Kárpáti Z.,
Terpó A. (1971) a Malus sylvestrist a tölgyesek, ezen belül a
mezofil tölgyesek között a gyertyánosok es nedvesebb talajú tölgyesek csoportjába
sorolja; ill. az Alföldön (rendszerint folyók mentén, lápos területeken) emlékeznek
meg felhasználhatóságáról "Alkalmazott növényföldrajz" c. munkájukban.
Ekképp utalnak azokra a termőhelyekre, ahol az ismételt – mesterséges – elterjesztés
siker reményében megpróbálható lenne.
A már korábbiakban
közölt társulásokat mintázó erdeinkbe – különösen a gyakori és közepes előfordulások
esetében – alkalmazható a vadalma szálankénti, ritka hálózatban történő elegyítése
a felújítás során. A fafajok megítélésében néhány éve jelentkező paradigmaváltás
hazánkban is megkönnyíti, hogy az erdőgazdálkodás partnerként viszonyuljon a
génmegőrzés munkáihoz, és annak eredményeit szükséges és hasznos segédeszközöknek
tekintse e cél szakszerû megvalósításában is.
Felhasznált Irodalom
- Andrásfalvy B. (1976):
A Duna mente népének gazdálkodása Tolna és Baranya megyében az ármentesítés
befejezéséig, Tanulmányok Tolna megye történetéből, Tolna Megyei Tanács Levéltára,
Szekszárd
- Bartha D., Kevey B.,
Morschauser T., Pócs T. (1995): Hazai erdőtársulásaink, in.: TILIA I. Szerk.:
Szmorad F., Tímár G., Erdészeti és Faipari Egyetem, Sopron
- Buttenschon, R. M., Buttenschon,
J. (1998): Population dynamics of Malus sylvestris stands in grazed and ungrazed,
semi-natural grasslands and fragmented woodlands in Mols Bjerge, Denmark,
Ann. Bot. Fennici 35 (4): 233-246. p.
- Csapody L, Csapody V.,
Rott F. (1966): Erdei fák és cserjék, Országos Erdészeti Főigazgatóság, Budapest
- Fr. Fiala, L. J. (1994):
Flowering crabapples - The Genus Malus, Timber Press, Portland, Oregon
- Gencsi L., Vancsura R.
(1992): Dendrológia, Mezőgazda Kiadó Budapest
- Hoffmann, H (1993): Zur
Verbreitung und Ökologie der Wildbirne (Pyrus communis L.) in Süd-Niedersachsen
und Nordhessen sowie ihrer Abgrenzung von verwilderten Kulturbirnen (Pyrus
domestica MED.) Mitt. Dtsch. Dendrol. Ges. 81, 27-71. p.
- Janb en, A., Walter,
P. (1997): Erhaltung von Wildapfel und Wildbirne in Hessen, AFZ / Der Wald
Nr. 11.: 602-604. p.
- Kárpáti Z., Terpó A.
(1971): Alkalmazott növényföldrajz, Mezőgazdasági Kiadó, Budapest
- Kleinschmit, J. (1998):
Erhaltung und Nutzung wertvoller Edellaubbaumarten, Forst und Holz Nr. 17.:
515-519. p.
- Kleinschmit, J., Wagner,
L (1995): Erhaltung von Wildobst in Nordwestdeutschland, AFZ / Der Wald Nr.
26.: 1458-1462. p.
- Kohnert, H. (1991): Neue
Möglichkeiten bei der heterovegetativen Vermehrung von Waldbaumen durch die
Chip-Veredlung, Die Holzzucht Nr. 41.: 30-32. p.
- Mátyás Cs., Bach I.,
Bordács S. [szerk.] (1998): Az erdei fás növények génmegőrzési alapelveinek
kidolgozása (az erdészeti genetikai erőforrások megőrzése c. nemzeti program
vázlata)”, OMMI, Budapest, 97 p.
- Schmidt, J. (1989):Die
Pflanzenanzucht von Sorbus-Arten und Wildobst, Forsttechnische Information
Nr. 5. 38-39. p.
- Schuman, K (1989): Obstgehölze
und Nuâbaume - die vergessenen Baumarten, AFZ Nr. 3839.: 1036-1039. p.
- Wagner, I. (1995): Identifikation
von Wildapfel (Malus sylvestris (L.) Mill.) und Wildbirne (Pyrus pyraster
(L.) Burgsd.), Voraussetzung zur Generhaltung des einheimischen Wildobstes,
Forstarchiv Nr. 6.: 39-47. p.
vissza